


Ahnenforschung zur Trockentoleranz
Prototyp der Schließzellregulation bei Moos gefunden
Pflanzen geben Wasser in Gasform über Spaltöffnungen in den Blättern an die Umgebungsluft ab. (Bildquelle: © Torsten Klemm / pixelio.de)
Landpflanzen haben die Fähigkeit, Kohlendioxid aus der Umgebungsluft zu binden. Bei diesem Prozess, die Photosynthese, entsteht Sauerstoff. Das Gas Kohlendioxid gelangt durch winzige Öffnungen in die Pflanzen. Im Gegenzug entweicht Feuchtigkeit aus diesen. Das Finetuning bei der Regulation von Gasaufnahme und Flüssigkeitsabgabe leisten bei höheren Pflanzen die Schließzellen. Auf der Suche nach dem Ursprung dieser spezialisierten Zellen sind Forscher nun ein Stück näher gekommen. Bereits die entwicklungsgeschichtlich sehr alten Grünalgen und Moose nutzen Enzyme und Ionenkanäle, die auch für die Regulation von Schließzellen notwendig sind. Erst bei den Laubmoosen wurden diese im Kontext der Regulation des Wasserhaushalts aktiv.
Der Klimawandel führt zu höheren Temperaturen und geringeren Niederschlägen. Um unsere Kulturpflanzen besser an die sich verändernden Umweltbedingungen anzupassen, müssen wir die Regulation des Wasserhaushaltes der Pflanzen zuerst verstehen. Landpflanzen sind ungemein wichtig für unseren Planeten, da sie Sauerstoff (O) erzeugen. Sauerstoff entsteht durch Photosynthese in den Chloroplasten der grünen Pflanzenteile, meistens sind das die Blätter. Bei diesem Prozess wird mit Hilfe der Sonnenenergie aus Wasser und Kohlendioxid ein Speichermolekül (Zucker) und Sauerstoff gebildet. Das Wasser und mit diesem die Nährstoffe werden über die Wurzeln aus dem Boden aufgenommen. Die Aufnahme von Kohlendioxid erfolgt aus der Luft.

In den Schließzellen des Laubmooses Physcomitrella patens befinden sich die ersten funktionsfähigen Kanalproteine, welche durch Trockenstress aktiviert werden können.
Bildquelle: © Pirex/ wikimedia.org/ CC0
Vor circa 500 Millionen Jahren eroberten die ersten Lebewesen, die Grünalgen, das Land. Sie waren nun nicht mehr andauernd von Wasser umgeben und in Dürreperioden sind sie vertrocknet. Eine Weiterentwicklung der ursprünglichen landbewohnenden Pflanzen sind die Moose. Sie haben eine Toleranz gegen Austrocknung entwickelt: Bei Wassermangel trocknen Moose langsam ein, bekommen sie wieder Wasser, können sie wieder Photosynthese betreiben und wachsen weiter. Die höher entwickelten Pflanzen können das nicht. Sie haben andere Mechanismen entwickelt, um den für die Photosynthese notwendigen Austausch mit der Umgebungsluft und den Schutz vor dem Austrocknen zu regulieren.
Spaltöffnungen regulieren den Wasserhaushalt
Die äußerste Schicht einer Gefäßpflanze, Epidermis genannt, hat besonders dicke Zellwände, die für die Stabilität des Organismus sorgen. Die aufgelagerten Wachsschichten der Cuticula sind fast undurchlässig für Wasserdampf. Damit wird die Transpiration an der Epidermis vermindert und die Pflanze vor Wasserverlust geschützt. Pflanzen haben aber meistens an der Blattunterseite winzige Öffnungen, die sogenannten Stomata oder auch Spaltöffnungen. Die Spaltöffnung hat im Aussehen die Form eines Schwimmreifens und besteht aus zwei bohnenförmigen Schließzellen, welche zusammen eine Pore bilden. Die angrenzenden Zellen besitzen Ionenkanäle, die bei Bedarf Salze in die Schließzellen pumpen. Erhöht sich die Salzkonzentration in einer Schließzelle, dann strömt Wasser passiv durch Osmose nach. Dadurch verändert sich der Zelldruck (Turgor) und die Pore der Spaltöffnung öffnet sich.
Spaltöffnungen sind also verschließbare Poren, die das luftgefüllte Adersystem der Pflanze mit der Außenluft verbindet. Sind sie geöffnet, erlauben sie den Austausch von gasförmigen Stoffen. Kohlendioxid gelangt in die Pflanze, Sauerstoff und Wasserdampf werden an die Umgebungsluft abgegeben. Sind die Stomata geschlossen, verhindern sie Wasserverlust zum Beispiel bei Trockenheit.
Pflanzen tauschen Kohlendioxid, Sauerstoff und Wasserdampf über Spaltöffnungen (Stomata) in ihren Blättern aus. Die molekulare Grundlage für die Regulation des Schließmechanismus ist jetzt bekannt.
Bildquelle: (Bildquelle: © iStock.com/ defun)
Schlüsselrolle des Enzyms OST1
Eine Wissenschaftlergruppe der Universität Würzburg hat in den vergangenen Jahren die Funktionsweise von Schließzellen untersucht. Entwicklungsgeschichtlich frühe Landpflanzen wie Moose können schon wie auch höherentwickelte Pflanzen bei Trockenheit das Stresshormon Abscisinsäure (ABA) bilden. ABA wiederum aktiviert die Produktion von Proteinen, welche die Moose vor dem Austrocknen schützen. Diese Schutzproteine werden Dehydrine genannt. Das Enzym OST1 (engl. Open STomata 1) ist maßgeblich an der Bildung der Dehydrine beteiligt.
Einige Moose (Laubmoose und Hornmoose) besitzen aber zudem auch schon Spaltöffnungen zur Regulation des Wasserhaushalts. Lebermoose hingegen besitzen keine Stomata. Schließzellen reagieren auf niedrige Luftfeuchtigkeit. Sie bilden dann das Stresshormon ABA. Durch ABA werden Salze mittels der Ionenkanäle aus den Schließzellen herausbefördert.Zusammen mit den Ionen verliert die Zelle Wasser, es kommt zur Verminderung des Zelldrucks. Die Schließzellen werden schlapp und schließen sich.
Die Forschergruppe aus Würzburg unter der Leitung von Professor Rainer Hedrich hat Pflanzen untersucht, bei denen dieser Schließmechanismus gestört ist. Sie haben Mutanten in der Modellpflanze Arabidopsis thaliana erzeugt, bei denen der Anionenkanal SLAC1 (engl. SLow Anion Channel 1) fehlt. Ohne dieses Kanalprotein kann die Spaltöffnung nicht geschlossen werden. Sie haben auch herausgefunden, dass die Proteinkinase OST1 nicht nur bei der Bildung der Austrocknungs-Schutzproteinen, die Dehydrine, eine Schlüsselfunktion hat. Die Anionenkanäle reagieren nicht nur auf ein bestimmtes Signal, sondern auf verschiedene Signale. Sie bekommen an unterschiedlichen Stellen von verschiedenen Proteinen Phosphat-Moleküle angeheftet und werden durch diese Signale aktiviert. Diesen chemischen Prozess nennt man Phosphorylierung. Fehlt jedoch die Proteinkinase OST1, reagieren die Schließzellen nicht mehr auf das Hormon ABA. Das Enzym hat also auch in der Aktivierung des Schließmechanismus der Stomata eine Schlüsselfunktion.
Es konnte gezeigt werden, wie die Signalkette abläuft: Trockenstress führt zur Bildung des Pflanzenhormons ABA. ABA aktiviert die Proteinkinase OST1, welche die Dehydrin-Bildung ankurbelt und gleichzeitig die SLAC1-Kanäle in den Schließzellen durch Phosphylierung anschaltet. Die Kanäle pumpen dann Ionen aus den Schließzellen, die Spaltöffnung wird geschlossen.
Was war zuerst da: Schutzproteine oder Kanäle?
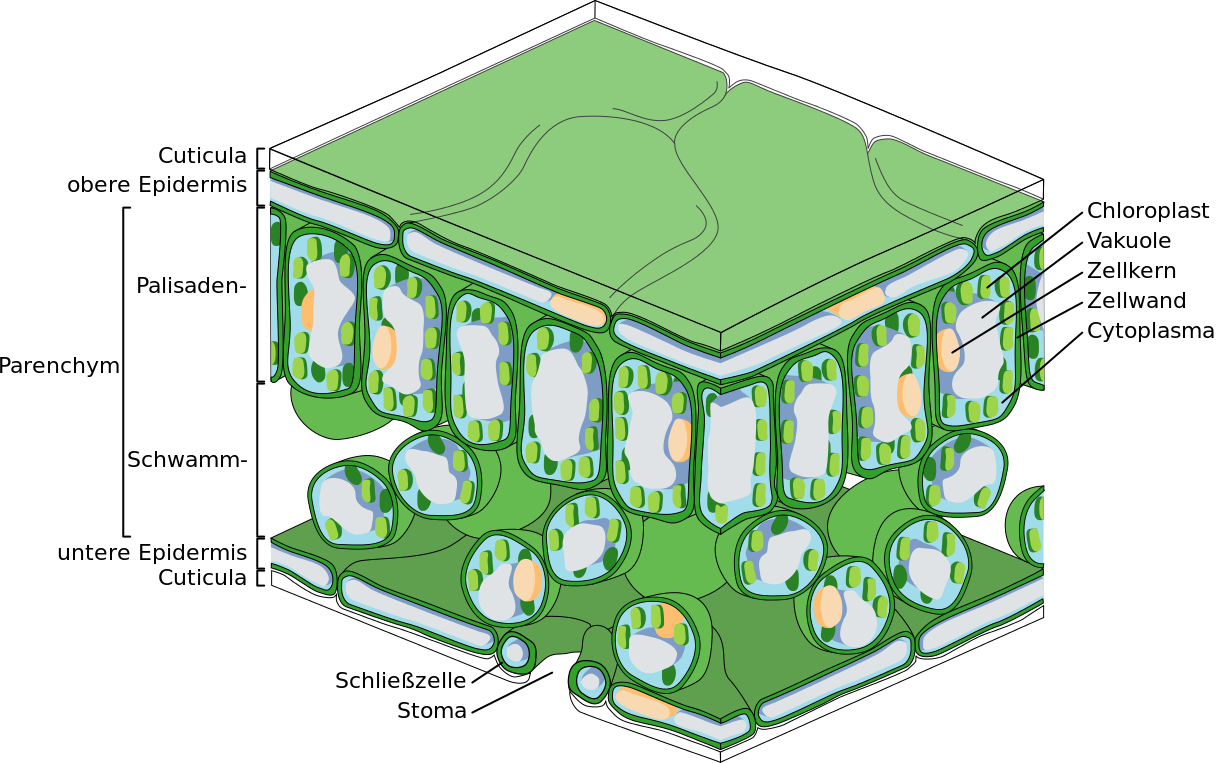
Der Aufbau eines Blattes im Querschnitt. Die schützende Wachsschicht, die Cuticula, ist ganz oben zu erkennen, während sich die Stomata ganz unten befinden.
Bildquelle: © Zephyris/ wikimedia.org/ CC BY-SA 3.0
Ein international zusammengesetztes Forschungs-Team ging der Frage nach, wann in der Stammesgeschichte der Pflanzen die Proteinkinase OST1 anfing, die Bildung der Schutzproteine zu steuern. Die Wissenschaftler aus Deutschland, Schweden, Spanien, Saudi-Arabien, Japan und den USA interessierte auch, ab wann in der Evolution dasselbe Enzym begann die Anionenkanäle zum Schließen der Spaltöffnungs-Pore anzuschalten. Sie isolierten die Gene für OST1 und SLAC1 aus vier verschiedenen Landpflanzen, um ihre Funktion zu untersuchen. Eine ursprüngliche Landpflanze stellt die mehrzellige Grünalge Klebsormidium nitens dar. Sie besitzt genauso wie das Lebermoos Marchantia polymorpha keine Spaltöffnungen. Erst das Moos Physcomitrella patens besitzt Spaltöffnungen auf der Sporenkapsel des Fortpflanzungsapparates.
Diese drei Pflanzen werden mit der Modellpflanze der Pflanzenforschung, die Ackerschmalwand (Arabidopsis thaliana), verglichen. Der Sequenzvergleich der OST1-kodierenden Gene zeigt, dass die Gene aus allen vier Pflanzen sehr ähnlich sind. Das bedeutet, sie besitzen ein gemeinsames Gen als Vorfahr, von dem sie abstammen. Die Wissenschaftler haben eine mutierte Arabidopsis-Pflanze erzeugt, welche keine Dehydrine mehr produzieren kann. Werden OST1-Genen aus der Alge und der beiden Moose in der Arabisdopsis-Mutante exprimiert, können bei Trockenstress wieder Dehydrin-Schutzproteine gebildet werden. Man kann also vermuten, dass die durch das Pflanzenhormon ABA induzierte Kaskade stammesgeschichtlich älter ist, als die durch das Enzym OST1 gesteuerte Aktivierung der Anionenkanäle in den Schließzellen.
Ionenkanal erst bei Laubmoos aktiv
Trotzdem besitzt auch die Grünalge schon ein Gen, das dem Ionenkanal SLAC1 ähnlich ist. Es sollte nun geklärt werden, ob diese Proteine auch schon funktionstüchtig sind. Die Anionenkanäle der Grünalge, der beiden Moose und der Pflanze Arabidopsis thaliana sind Membranproteine. Die Wissenschaftler benutzten die Eizellen des Krallenfrosches Xenopus laevis als Modell, um die Funktion dieser Membranproteine zu untersuchen. Die Proteinkinase OST1-Analoge aller vier Organismen können den Anionenkanal der Modellpflanze Arabidopsis aktivieren. Jedoch nur das höher entwickelte Laubmoos Physcomitrella bildet ein durch ABA aktivierbares Proteinkinase-Anionenkanal-System aus, welches mit dem der Schließzellen der Arabidopsis-Pflanze vergleichbar ist.
Sukkulenten behalten auch bei großen Hitzen einen kühlen Kopf. Hilfreich ist dabei die helle Cuticula, welche einen Großteil der Sonnenstrahlen reflektiert.
Bildquelle: © gnubier/ pixelio.de
Die Wissenschaftler vermuteten, dass die Gene für die Anionenkanäle SLAC1 sich während der Weiterentwicklung der Landpflanzen erst verändert haben, so dass eine Aktivierung durch die Proteinkinase OST1 ermöglicht wurde. Bei Sequenzvergleichen der Anionenkanal-Gene fällt auf, dass weder die Grünalge noch das Lebermoos die Phosphorylierungsstellen besitzen, welche für die Aktivierung durch Proteinkinasen notwendig sind. Beim schwach aktiven Anionenkanal des Laubmooses wurden Sequenzen durch entsprechende Abschnitte aus dem Arabidopsis-Gen ausgetauscht. Der rekombinante Ionenkanal kann nun durch alle Proteinkinasen OST1 der vier untersuchten Arten aktiviert werden, was vorher nicht möglich war.
Diese Erkenntnisse decken die entwicklungsgeschichtliche Entstehung des Wasserspar-Mechanismus noch nicht vollständig auf. Die Schließzellen von Laubmoosen reagieren, wenn überhaupt, nur schwach auf das Wasserstresshormon Abszisinsäure (ABA). Weitere Untersuchungen sollen an Farnen und anderen stammesgeschichtlich frühen Gefäßpflanzen vorgenommen werden, um Klarheit zu schaffen.
Ziel der Forscher ist es, mit Hilfe ihrer Erkenntnisse Kanalproteine so zu verändern, damit Kulturpflanzen Trockenstress auch in Zeiten des Klimawandels besser verkraften und stabile und sichere Erträge liefern.
Quelle: C. Lind et al. (2015): Stomatal Guard Cells Co-opted an Ancient ABA-Dependent Desiccation Survival System to Regulate Stomatal Closure. In: Current Biology Vol. 25, S. 928-935, (30. März 2015), doi.org/10.1016/j.cub.2015.01.067
Zum Weiterlesen auf Pflanzenforschung.de:
- Wie Pflanzen Wasser sparen
- Kompromiss bei Trockenheit: Stomata nur so weit schließen, wie nötig
- An- und Ausschalter für Stomata entdeckt
- Stoffwechselweg entdeckt, Wie Pflanzen die Anzahl ihrer Stomata regulieren
- Geringe Luftfeuchtigkeit gibt das Signal zum Abschotten
Titelbild:Pflanzen geben Wasser in Gasform über Spaltöffnungen in den Blättern an die Umgebungsluft ab. (Bildquelle: © Torsten Klemm / pixelio.de)
