


Pflanzen-Pathogen-Interaktion

Pathogene schaden Pflanzen. (Quelle: © iStockphoto.com/ Keith Gentry)
Pflanzen sind einer Vielzahl von Feinden ausgesetzt. Neben großen Widersachern wie dem Menschen oder verschiedenen Tieren müssen Pflanzen auch mit kleinsten Lebewesen wie Viren oder Mikroorganismen fertig werden. Die Bezeichnung "Pflanzen-Pathogen-Interaktion" steht somit für ein Zusammenspiel zwischen einer Pflanze und einem Schaderreger.
Pflanzen haben in Jahrmillionen verschiedene Abwehrmechanismen entwickelt, mit denen sie sich gegen aktiv oder passiv eindringende Pathogene oder Fressfeinde (Pilze, Viren, Bakterien, Wirbellose, Insekten, Säugetiere) schützen können. Sie verfügen damit über ein ähnlich effektives ’Immunsystem’ wie Tiere und der Mensch, das aus mehreren Barrieren besteht. Obwohl Wildpopulationen in der Regel resistent sind, kommt es jedoch bei Nutzpflanzen immer wieder zu schweren Ernteausfällen, oft hervorgerufen durch einen einzigen Erreger (bekanntes Beispiel: Die Kartoffelfäule, hervorgerufen durch den Pilz Phytophthora infestans, in deren Folge über eine Million Menschen in Irland Mitte des 19. Jahrhunderts verhungerten). Alljährlich gehen etwa 30 Prozent der Ernte durch solche Pflanzenkrankheiten verloren. Daher wird in diesem Bereich intensiv geforscht.
Was ist Pflanzen-Pathogen-Interaktion?
Bei der Pflanzen-Pathogen-Interaktion handelt es sich um ein Zusammenspiel zwischen einer Pflanze und einem Schaderreger. Sie beginnt mit dem Versuch des Pathogens (Virus, Bakterium, Pilz), die Pflanze zu besiedeln. Auf dieses Eindringen von außen erfolgt eine Reaktion der Pflanze. Wildpflanzen werden für gewöhnlich gut mit eindringenden Schädlingen fertig, da sie umfangreiche Abwehrmechanismen entwickelt haben, mit denen sie sich gegen die meisten potentiellen Schädlinge zur Wehr setzen können. Diese Grundresistenz nennt man auch Basisresistenz, Basis-Inkompatibilität oder Nichtwirtsresistenz.
Sie ist in allen Pflanzenfamilien auf die eine oder andere Art vorhanden und bewirkt, dass viele Pathogene die Pflanze nicht als Wirt erkennen können oder durch bestimmte unspezifische (generell auf Eindringlinge ausgelegte) Maßnahmen von vorne herein abgeblockt werden. ’Erkannt’ werden die eindringenden Pathogene durch sogenannte generelle Elicitoren, also bestimmte Komponenten, die in allen der Pflanze bekannten Pathogenen vorkommen. Die Abwehr, die sie bewirken, wird als unspezifisch oder horizontal bezeichnet, weil sie in die Breite wirkt und nicht nur auf einen speziellen Pathogen.

Wurzeln sind ein Einfallstor für Pathogene.
Bildquelle: © iStockphoto.com/ carroteater
Wirtsspezifische Resistenzen
Problematisch wird es für die Pflanze, wenn der Pathogen einen Weg gefunden hat, die generellen Abwehrmechanismen zu umgehen. Aus der Basis-Inkompatibilität wird jetzt eine Basis-Kompatibilität oder Wirtssuszeptibilität (Umgangssprachlich wird Suszeptilität mit Empfänglichkeit, Anfälligkeit oder Empfindlichkeit umschrieben). Dagegen entwickelt die Pflanze spezifische Abwehrmechanismen, die konkret auf diesen einen Schädling zielen (rassespezifische Wirtsresistenz). Diese Resistenz ist genau auf einen Pathogen spezialisiert und sehr wirkungsvoll (vertikale Resistenz).
Sie endet oft mit der sogenannten Hypersensitiven Reaktion (HR), dem gesteuerten Absterben befallener Pflanzenzellen. Die Resistenz kann aber auch leicht außer Gefecht gesetzt werden, etwa wenn sich beispielsweise das Genom des Schädlings ändert und die Pflanze den Feind nicht mehr erkennen kann. Gerade hochspezialisierte Nutzpflanzen sind auf eine spezifische Resistenz ausgelegt und haben einem veränderten Schaderreger nichts entgegenzusetzen, so dass es für den Erreger einfach ist, die Verteidigung der Pflanze zu umgehen und sie zu besiedeln.
Gen-für-Gen-Hypothese
Die Beschreibung der wirtsspezifischen Resistenz wird über die Gen-für-Gen-Hypothese (Flor 1971) beschrieben. Diese Hypothese benennt die virulenten Gene als Avirulenzgene (AVR) des Pathogens und die daran angepassten, komplementären Resistenzgene (R) der Pflanze. Existiert ein R zum AVR, bleibt die Pflanze gesund, ist also resistent. Fehlt das passende R, wird die Pflanze krank.
Guard-Hypothese
Die Guard-Hypothese verfeinert die Gen-für-Gen-Hypothese. Es interagieren die über die AVR-Gene codierten Proteine zuerst als Virulenzfaktoren mit bestimmten Proteinen (Virulenz-Targets) der Wirtspflanze. Sie bilden sogenannte Virulenz-Target-AVR-Komplexe, über die der Pathogen versucht, den Stoffwechsel der Wirtszelle für sich zu manipulieren. Diese Interaktion oder Störung des Ablaufs der Wirtszelle wirkt als spezifischer Elicitor und wird von den R-Proteinen (guards) wahrgenommen. Diese leiten dann die Gegenreaktion ein.
Wie erfolgt die Besiedlung einer Pflanze durch einen Schaderreger?
Klassische Einfallstore für Schaderreger in eine Pflanze sind Risse oder Beschädigungen in der schützenden Wachsschicht (Cuticula) oder einfach die Spaltöffnungen (Stomata) an der Blattunterseite. In der Regel versuchen Pathogene über diesen Weg, bestimmte Substanzen in die Pflanzenzelle zu bringen, die ihnen die Invasion erleichtern. Bakterien versuchen z. B. mehrere Proteine (Effektoren) direkt ins Innere der Pflanzenzelle zu spritzen und so die Immunreaktion der Pflanze zu blockieren (Type-III-Secretion-System, TTSS). Pilze versuchen zunächst, die Zellwände aufzuweichen, damit sie in die lebenden Zellen eindringen können. Sie verwenden dafür beispielsweise spezielle Polygalacturonasen. Anschließend bilden sie spezielle Saugorgane, sogenannte Haustorien, um der Wirtszelle Nährstoffe abzuziehen. Auch sie nutzen Effektoren, allerdings gibt es über die Infektionswege bisher nur wenige Erkenntnisse.
Durch das evolutiv in vielen Jahrmillionen gewachsene Schädlingsabwehr-System ist es für potentielle Erreger sehr schwer, eine Pflanze zu besiedeln. Sie müssen dazu entweder vermeiden, dass einer der diversen Rezeptoren sie erkennt oder sie müssen die Abwehrmaßnahmen der Pflanze außer Kraft setzen.
Wie wehren sich Pflanzen gegen Schädlinge?
Im Unterschied zum Immunsystem von Tieren, das zentral organisiert ist und spezielle Abwehrzellen besitzt, reagieren bei Pflanzen die einzelnen Zellen auf das Eindringen von außen. Trotzdem gibt es einige Ähnlichkeiten zwischen dem pflanzlichen und tierischen Immunsystem, beispielsweise beim programmierten Zelltod. Diese Ähnlichkeiten legen nahe, dass die Ursprünge beider Systeme gleich sind.
Das Abwehrsystem der Pflanzen besteht aus zwei Stufen. Zunächst bremsen die generell vorhandenen aktiven und passiven Schutzschirme (präformierte Barrieren) einen eindringenden Schädling aus. Falls doch einer durchkommt, muss er anschließend die durch sein Eindringen in Alarm versetzten aktiven Verteidigungsmaßnahmen (induzierte Abwehr) entweder umgehen oder außer Gefecht setzen. Diese Abwehr ist wiederum in zwei Bereiche aufgeteilt, einmal einen ’äußeren Radar’ auf der Zelloberfläche und einen ’inneren Radar’ im Inneren der Zelle.
Präformierte Barrieren
Sie stellen die erste Stufe der pflanzlichen Abwehr dar. Formen der passiven Abwehr von Pathogenen sind in der Pflanze generell vorhanden, müssen also nicht erst nach dem Eindringen eines Pathogens gebildet werden. Hier wirkt zunächst die äußere Wachsschicht der Blätter (Cuticula) als Stopper vor allem von Pathogenen, die auf Feuchtigkeit angewiesen sind (Pilze). Auch ist die Oberfläche eines unverletzten Blattes dadurch versiegelt, bildet also eine nahezu undurchdringliche Barriere. Weitere Schutzmechanismen sind Haare auf der Blattoberfläche oder zum Schutz von Spaltöffnungen (werden von Pilzen gerne als Einfallstor genutzt).
Sekundäre Pflanzenstoffe
Weitere unspezifische, aber aktive Barrieren bestehen aus bestimmten chemischen Verbindungen (sogenannte konstitutive Pflanzenabwehrstoffe oder Pre-Infectional Compounds), die entweder die eindringenden Hyphen direkt angreifen (hydrolytische Enzyme) oder deren Enzyme blockieren. Diese Enzyminhibitoren (sogenannte Polygalacturonasen-inhibierenden Proteine oder PGIPs ) stoppen Enzyme, die der Pilz in die Zelle bringt, um die Zellwand aufzuweichen und sich besser verbreiten zu können. Die Inhibitoren befinden sich in der Zellwand.
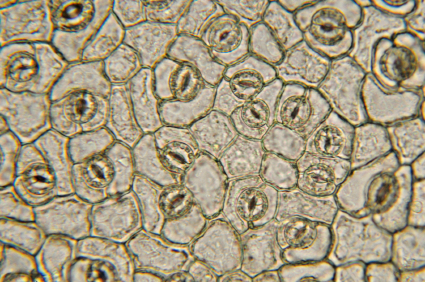
Zellstruktur von Pflanzenzellen. Gut zu erkennen sind Zellwände und Spaltöffnungen. Während die Zellwand zur Abwehr von Pathogenen verstärkt werden kann, sind Spaltöffnungen häufig ein Einfallsort für Pathogene.
Bildquelle: © iStockphoto.com/ Nancy Nehring
Andere chemische Verbindungen (sogenannte Phytoanticipine) lagern in der Vakuole, die durch einen Angriff durch einen Pathogen zerstört wird, wodurch die Substanzen freigesetzt werden. Sie bestehen z. B. aus cyanogenen Glycosiden (werden nach Angriff von ß-Glucosidasen umgewandelt in sogenannte Aglycone, ein weiterer Schritt setzt giftige Blausäure frei), Glucosinolaten (Senföle, die Kreuzblütler zur Pilzabwehr einsetzen), Saponinen (reagieren mit Bestandteilen der Pilzzellwand und machen sie porös). Andere Stoffe sind beispielsweise Phenole, Lactone.
Induzierte Abwehr
Nachdem der Erreger die präformierten Barrieren überwunden hat, muss er sich mit einer durch sein Eindringen aktivierten Abwehr auseinander setzen. Hierbei handelt es sich um eine Reaktion, die erst nach der Erkennung eines Pathogens eintritt, sie ist die zweite Stufe der pflanzlichen Abwehr. Sie wird in zwei Komponenten unterteilt: Im ersten Schritt erfolgt die Warnung vor ’Feinden’ durch spezielle Rezeptoren (sogenannte Pattern Recognition Receptors oder PRRs) die auf der Zelloberfläche sitzen und bestimmte Komponenten erkennen, die nur in Pathogenen vorkommen.
Diese Komponenten werden als Elicitoren (Botenstoffe) oder PAMPs (Pathogen-Associated Molecular Patterns) bezeichnet. Bekannte PAMPs sind z. B. PEP13, ein Peptid-Fragment aus Phytophthora sojae, bakterielles Flagellin (flg 22) oder Chitin aus Pilzzellwänden. Ihre Erkennung leitet die Produktion von unspezifisch wirksamen Stoffen ein (PAMP-Triggered-Immunity oder PTI).
PRRs bestehen in der Regel aus einer extrazellulären, leucin-reichen ’Antenne’ (Leucin-Rich-Repeat-domain, LRR) und einer intrazellulären Kinase. Bei Kontakt mit einem PAMP bildet das PRR einen Komplex mit einer BAK1 (Brassinosteroid-Insensitive 1-Associated Kinase 1). Dieser aktivierte Komplex löst zum Beispiel durch Vermittlung einer BIK1 (Botrytis-Induced Kinase 1) oder MAPK1 (Mitogen-Activated Protein Kinase) ein Signal aus, das die weitere Immunreaktion bewirkt.
Nachdem die Wahrnehmung von Feinden auf der Zelloberfläche durch PAMPs möglich ist, besitzt die Zelle noch ein zweites, hochspezifisches Warnsystem im Inneren der Zelle. Hier sind es ebenfalls pathogene Bestandteile (Effektoren) die den Alarm auslösen. Das Warnsystem überwacht die wichtigen Enzyme und registriert Veränderungen, die durch eindringende Effektoren bewirkt werden (Effector-Induced-Immunity oder ETI). Es hat ein NB-LRR-Protein, bestehend aus einer Bindungsdomäne (Nuclear Binding domain, NB) sowie der LRR und der dazugehörigen Kinase und löst eine ähnliche Abwehr aus wie die PRRs. Da aber die Bandbreite der Effektoren größer ist und sie sich auch schneller verändern können, reagiert dieses System spezifischer und deutlich schneller als das PTI. Es kann auch einen programmierten Zelltod bewirken.
Als Antwort auf dieses hochsensible Abwehrsystem haben viele Pathogene Effektoren entwickelt, die als sogenannte Suppressoren fungieren. Sie sind vom Pathogen entwickelte Komponenten, die die PTI unwirksam machen sollen, so dass die Pflanze die Besiedelung durch einen Pathogen nicht ’bemerkt’.
Folgen der induzierten Abwehr in der Pflanzenzelle
Verstärkung von Zellwänden
Hierzu wird rund um die Verwundung zwischen Zellwand und Plasmamembran Callose oder Lignin eingelagert und somit die Zellwand verstärkt. Das verlangsamt oder stoppt den Pilz, er wird quasi ’eingemauert’, so dass er nicht mehr weiter vordringen kann. Lignin-Vorstufen haben außerdem eine toxische Wirkung auf Pathogene.
Oxidative Burst
Er wird bewirkt durch die Aktivierung von speziellen Ezymen, die Sauerstoff-Radikale freisetzen, zum Beispiel der in der Zellwand ansässigen NAPDH-Oxidase, aus deren Reaktion mit Luftsauerstoff Hydroxyl-Radikale (OH*), Superoxid-Radikale (O2*) und Wasserstoffperoxid (H2O2), sogenannte Radical Oxidative Species, (ROS) entstehen. Die stark oxidierende Wirkung dieser Moleküle bewirkt eine Inaktivierung von Enzymen, ein Aufoxidieren von Lipiden in der Zellwand sowie Abbau von Nukleinsäuren sowohl bei der betroffenen Pflanzenzelle als auch beim Eindringling.
Außerdem ist sie für eine Auslösung einer ganzen Reihe von Immunreaktionen verantwortlich, wie zum Beispiel der Quervernetzung von Zellwandproteinen, der Hypersensitiven Reaktion sowie der Produktion von antimikrobiellen Stoffen. Diese Abwehrreaktion gehört zu den schnellsten und kann innerhalb von Sekunden greifen. Auch das um einen Infektionsherd gehäuft auftretende Stickstoffmonoxid (NO) spielt anscheinend eine wichtige Rolle bei der Weitervermittlung von Signalen, die zur Auslösung der Immunabwehr führen, indem es zum Beispiel wichtige Abwehrgene aktiviert.
Hypersensitive Reaktion
Die hypersensitive Reaktion ist ein Zusammenspiel aus verschiedenen Abwehrreaktionen, die zu einem von der Pflanze kontrollierten Absterben befallener Zellen führt und dadurch den Pathogenen eingrenzt oder stoppt. Ausgelöst wird das Absterben beispielsweise durch einen Oxidative Burst oder durch eine Überproduktion von bestimmten Stoffen, die die Zelle abtöten. Das Absterben der Pflanzenzellen in einem eng begrenzten Bereich um den Pathogen (programmierter Zelltod) bewirkt, dass diesem Wasser und Nährstoffe entzogen werden und er zusammen mit der Zelle eingeht. Das Absterben wird äußerlich als dunkler bis schwarzer Fleck (Nekrose) sichtbar. Gleichzeitig werden von den absterbenden Zellen Signale an die benachbarten Zellen weiter gegeben, die für eine Verstärkung von Zellwänden durch Einlagerung von Lignin, Callose oder Querwandvernetzungen sorgen.
Systemische Resistenzen (systemic aquired resistance, SAR)
Durch den Befall mit einem Pathogen können auch an den nicht befallenen Stellen Resistenzen entstehen (systemisch erworbene Resistenzen). Diese werden wiederum durch Signalstoffe bewirkt, die bei Befall ins Phloem abgegeben werden, sich auf diesem Weg über die ganze Pflanze verteilen und sie in Alarmbereitschaft versetzen. Eine wichtige Rolle spielen hierbei Jasmonsäure, Ethylen und Salicylsäure. Manche dieser Signalstoffe sind flüchtig (etwa methylierte Salicylsäure), so dass sie auch benachbarte Pflanzen warnen können. Als Folge der systemischen Resistenz werden vermehrt antimikrobielle Stoffe gebildet.
Bereits gegen Ende der 70er Jahre konnte gezeigt werden, dass eine Begasung von Tabak (Nicotiana tabacum) mit Salicylsäure zu einer erhöhten Resistenz gegenüber dem Tabakmosaikvirus führt.
Pathogeninduzierte Proteine (pathogen-related proteins, PR)
Sie bezeichnen Proteine, die Bakterien und Pilze angreifen können, zum Beispiel Glucanasen oder Chitinasen, die die Zellwände von Pathogenen abbauen. Sie sind nur im Bereich befallener Pflanzenteile zu finden.
Phytoalexine
Phytoalexine bezeichnen Stoffe, die durch Genaktivierung ’de novo’ produziert werden, sobald ein Eindringen eines Pathogens vorliegt. Die Substanzen (Terpene, Flavonoide, Cumarine, Alkaloide) sind etwa 24 Stunden nach Infektion nachweisbar und erreichen etwa drei Tage später ein Konzentrationsmaximum, allerdings nur einige Millimeter um die betroffene Stelle. In nicht befallenen Bereichen der Pflanze sind sie nicht nachweisbar.
Pflanzen können sich gegenseitig warnen
Ein bekanntes Beispiel für die Warnung vor Feinden ist die Schirmakazie (Acacia tortilis). Sobald Herbivoren (in diesem Fall Antilopen und Giraffen) anfangen, an einer Akazie zu fressen, wird durch Bestandteile des Speichels (als Elicitor) die Bildung des gasförmigen Phytohormons Ethen angeregt. Dieses löst in der angefressenen Akazie die Produktion von übel schmeckenden Gerbstoffen (Tanninen) aus und ’warnt’ gleichzeitig die benachbarten Akazien vor den Tieren, die ebenfalls mit der Produktion von Tanninen beginnen, so dass sämtliche Akazien in der Umgebung für die Tiere innerhalb kurzer Zeit ungenießbar werden. Allerdings haben sich die Tiere auf ihre Weise an diesen Abwehrmechanismus angepasst, so dass sie nur kurz an einer Akazie fressen und sich gegen den Wind weiterbewegen, um Akazien zu erreichen, die noch nicht von der Warnung ’Wind’ bekommen haben.
Die Pflanzenforschung sucht nach hoch effektiven Rezeptoren, um Pflanzenpathogene zu entdecken.
Bildquelle: © iStockphoto.com/ Andreas Reh
Forschung im Bereich Pflanzen-Pathogen-Interaktion
Aufgrund der hohen Ernteausfälle durch Pathogene wird intensiv daran geforscht, wie man eine verbesserte Resistenz in Nutzpflanzen bewirken kann. Dazu ist es zunächst notwendig zu verstehen, wie die verschiedenen Schaderreger die Pflanze angreifen, wie die Pflanze sie erkennt und sich ’wehrt’ und wie der Erreger wiederum diese Verteidigungslinien zu umgehen versucht. Intensive Forschungen dazu gibt es beispielsweise am Leibniz-Institut für Pflanzengenetik und Kulturpflanzenforschung (IPK) in Gatersleben (Patrick Schweizer) und am Max-Planck-Institut für Züchtungsforschung (MPIZ) in Köln (Paul Schulze Lefert, Jane Parker).
Die Pflanzenforschung zielt besonders auf die Entdeckung oder Entwicklung von hocheffektiven Rezeptoren, die möglichst viele pathogene Effektoren bzw. hoch-virulente Effektoren erkennen können. Dazu werden besonders die NB-LRR-Proteine erforscht sowie die Möglichkeiten, sie in die Pflanzen zu bringen, entweder durch Züchtung oder über gentechnische Methoden. Wie bereits beschrieben sind NB-LRR-Proteine eine Kombination aus Bindungsdomäne (Nuclear Binding Domain), langen leucinreichen Antennen (Leucin-Rich-Repeat-Domain) sowie der dazugehörigen Kinasen.
Auch die Gensequenzen verschiedener Pathogene werden erforscht, um möglichst viele Effektoren zu finden, mit denen dann wiederum Rezeptoren aus Wildpflanzen getestet werden können, um neue Schutzmöglichkeiten zu finden. Ein weiterer möglicher Weg zur verbesserten Resistenz von Pflanzen besteht im Austausch der PRRs verschiedener Pflanzenfamilien. Da Pathogene sich auf eine bestimmte Pflanzen(familie) spezialisieren, bereitet es ihnen einige Schwierigkeiten, bei der speziellen Familie auf ein unspezifisch reagierendes PTI aus einer anderen Pflanzenfamilie zu treffen, dass zwar generell Pathogene erkennt, bei dem aber das von diesem Pathogen genutzte Einfallstor fehlt. Diese Art von ’Wissenstransfer’ wurde bereits erfolgreich mit einem PRR von Arabidopsis (EF-Tu-Rezeptor) an Tomaten getestet (Lacombe et al. 2010).
Mehltau-Resistenz bei Gerste
Echter Mehltau (Blumeria graminis f.sp. hordei) tritt bei Gerste relativ häufig auf. Allerdings gibt es Gerstensorten, bei denen der Mehltau keine Chancen hat, die also immun sind. Forscher am Max-Planck-Institut für Züchtungsforschung haben entdeckt, dass diese Resistenz auf einem Defekt in einem Gen beruht. Dieses codiert für ein Membran-Protein, das der Pilz als Einfallstor benutzt, um die Gerste zu besiedeln. Ist dieses Protein falsch gebaut (durch das defekte Gen), kann der Pilz folglich nicht eindringen, weil er mit dem kaputten Protein nichts anfangen kann – wenn die Tür fehlt, kann man nicht eintreten.
Innere Immunabwehr
Pflanzen haben verschiedene Warnsysteme, die beim Eindringen eines Pathogens eine Warnung absetzen und eine Immunantwort auslösen. Diese Vorgänge sind Gegenstand intensiver Forschung, vor allem die Kommunikation zwischen dem Radar und der folgenden Immunreaktion. Am Max-Planck-Institut für Züchtungsforschung ist jetzt ein wichtiges Enzym entdeckt worden, das offenbar zwischen dem inneren Radar einer Pflanzenzelle (ETI) und der hochspezifischen Immunantwort vermittelt und den Notruf des inneren Radars auffängt. Das Protein EDS1 erhöht seine Präsenz im Zellkern und im Cytoplasma der befallenen Zelle, sobald der Radar Alarm schlägt. Dann scheint es bei der Produktion von Abwehrstoffen ’behilflich’ zu sein, auch wenn es offenbar nicht direkt an der Produktion beteiligt ist. Welche Funktion es genau erfüllt, ist noch unklar, aber fest steht: Erfüllt es seine Funktion nicht, ist die Immunantwort der Zelle schwach oder es wird sogar der programmierte Zelltod herbeigeführt.
Zum Weiterlesen und vertiefendes Wissen:
- Arbeit zum "Stress an der Gerste durch den Verursacher des Echten Mehltaus" der Universität Halle
- Begriff "Pflanzliche Abwehr von Pathogenen" auf Wikipedia
Links zu Forschungsbeispielen:
- Artikel zum Projekt GABI-PHENOME "Gerste soll sich vor Pilzbefall schützen können" auf Pflanzenforschung.de
- Interview zum Projekt GABI-PHENOME "Pilzresistenz der Gerste verstehen und ausbauen" auf Pflanzenforschung.de
- Artikel zum Projekt GABI-GRASP "Ein „gutes Tröpfchen“ noch besser machen – das ist das Ziel des GRASP-Forschungsprojektes" auf Pflanzenforschung.de
- Interview zum Projekt GABI-GRASP "Wie kann man Wein noch besser machen?" auf Pflanzenforschung.de
- Forschungsarbeit "Importierte Fitness - Kölner Max-Planck-Forscher entschlüsseln Resistenzmechanismus der Gerste gegen Pilzinfektionen" der Max-Planck-Gesellschaft Pflanzenzüchtungsforschung
- Forschungsarbeit "Auch Pflanzen haben eine doppelte Abwehrkette - Kölner Max-Planck-Forschern weisen nach, dass die dauerhafte Resistenz von Pflanzen gegen Pilzparasiten auf einem mehrstufigen Abwehrmechanismus beruht" der Max-Planck-Gesellschaft Pflanzenzüchtungsforschung
- Forschungsarbeit "Universeller Resistenzmechanismus bei Pflanzen entdeckt - Kölner Max-Planck-Forscher entdecken Anlagen für eine natürliche Mehltauresistenz bei der Modellpflanze Arabidopsis" der Max-Planck-Gesellschaft Pflanzenzüchtungsforschung
- Forschungsarbeit "Notfallplan in Pflanzenzellen - Max-Planck-Wissenschaftler decken ausgeklügeltes Abwehrsystem in Pflanzen auf" der Max-Planck-Gesellschaft Pflanzenzüchtungsforschung
- Forschungsarbeit "Krisenmanager der Pflanzenabwehr - Biologen haben einen wichtigen Helfer für eine vollständige Immunantwort bei Pflanzen entdeckt" der Max-Planck-Gesellschaft Pflanzenzüchtungsforschung
